



Justicieae
Lucinda A. McDade and Carrie Kiel





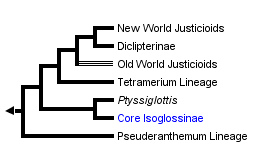

This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxThe phylogeny presented above is based on McDade et al. (2000), Kiel et al. (2006) and our on-going research.
Introduction
With more than 2000 species, Justicieae are the largest of the major lineages of Acanthaceae. The lineage is as widely distributed as the family as a whole and encompasses, arguably, as much diversity of plant morphological traits as well. The images presented above and on the pages for sublineages of Justicieae will give you some appreciation for this diversity.
Characteristics
McDade et al. (2000) proposed that tricolporate hexapseudocolpate pollen (see image below) is a synapomorphy for Jusiticieae: this trait occurs only in Justicieae although there have been many subsequent evolutionary changes in pollen morphology in the group.

Tricolporate hexapseudocolpate pollen grains. © 2007
Discussion of Phylogenetic Relationships
The work of McDade et al. (2000) provided strong support for monophyly of Justicieae and for many aspects of relationships within the lineage. The basal members of the group (the Pseuderanthemum lineage) retain the plesiomorphic character of having four staminal elements (i.e., either four fertile stamens or two fertile stamens and two staminodes). The Pseuderanthemum lineage was only weakly supported as monophyletic in the analysis of McDade et al. (2000), and work in progress suggests that there may be two or more basal lineages of Justicieae that retain the plesiomorphic staminal character.
Isoglossinae and the other more phylogenetically distal members of Justicieae have only two stamens, having lost even the staminodal vestiges of the other pair of stamens. The exception here is that some species of Ptyssiglottis are reported as having staminodes. Interestingly, the position of Ptyssiglottis as the basal member of Isoglossinae was not strongly supported in either tha analysis of McDade et al. (2000) or of Kiel et al. (2006). It seems that a more complete sample of Justicieae will be needed to place this genus with confidence.
The Tetramerium lineage is one of the most strongly supported clades of Acanthaceae to emerge from any of our analyses. It is remarkable that this clade has no known morphological synapomorphies although constituents can usually be recognized by presence of the synapomorphies of Justicieae as a whole (i.e., tricolporate hexapseudocolpate pollen) and of plants distal to the Pseuderanthemum lineage (i.e., complete loss of two stamens) and the absence of the synapomorphies that characterize 'neighboring' lineages (i.e., Isoglossinae: 'gürtelpollen'; rugula and anther complexity: 'justicioids'). We will shortly post here the results of our recently completed analysis of the Tetramerium lineage that included members of all recognized genera that have been associated with the group. One very interesting result of this study is that, among New World members of the lineage, hummingbird pollination has evolved at least seven times. Our results indicate that floral morphology can change extremely rapidly, evolutionarily speaking, in this group. Species that are quite closely related often differ dramatically in floral morphology.
The remaining members of Justicieae - at least 1100 species strong - are part of a strongly supported clade that we refer to informally as 'justicioids.' These plants share two synapomorphies: (1) the internal, upper surface of the corolla has a channel or 'rugula' in which the style lies at anthesis and (2) stamens have thecae that are not parallel, have various apical or basal appendages, and are not always both fertile (referred to by McDade et al. 2000 as 'anther complexity'). This last character actually represents several likely independent characters that deserve further study.
Old World 'justicioids' belong to several clades that are basal to New World 'justicioids' , Diclipterinae. That is, Old World 'justicioids' are not monophyletic but instead seem to form several lineages. A taxonomic complexity is that species treated in the large, nearly cosmopolitan genus Justicia occur in both the Old and New World 'justicioid' groups. This means that considerable changes of species names will be needed once the patterns of relationships are clear.
References
Kiel, C. A., L. A. McDade, T. F. Daniel, and D. Champluvier. 2006. Phylogenetic delimitation of Isoglossinae (Acanthaceae: Justicieae) and relationships among constituent genera. Taxon 55 (3): 683-694.
McDade, L. A., T. F. Daniel, S. E. Masta, and K. M. Riley. 2000. Phylogenetic relationships within the tribe Justicieae (Acanthaceae): evidence from molecular sequences, morphology, and cytology. Annals of the Missouri Botanical Garden 87: 435-458.
Title Illustrations

| Scientific Name | Anisacanthus thurberi A. Gray |
|---|---|
| Location | Arizona |
| Comments | Anisacanthus is a member of the Tetramerium Lineage of Justicieae |
| Specimen Condition | Live Specimen |
| Copyright | © 2003-2006 T. Beth Kinsey |

| Scientific Name | Hypoestes aristata (Vahl) Sol. ex Roem. & Schult. |
|---|---|
| Comments | Hypoestes is a member of Diclipterinae |
| Specimen Condition | Live Specimen |
| Copyright | © 2006 Zephyrus |

| Scientific Name | Ruttya ovata Harvey |
|---|---|
| Location | KwaZulu-Natal, South Africa |
| Comments | Ruttya is a member of the Pseuderanthemum lineage of Justicieae. |
| Specimen Condition | Live Specimen |
| Copyright |
© 2006

|

| Scientific Name | Justicia sp. L. |
|---|---|
| Comments | This species, and so far as is known all other New World species assigned to the genus Justicia, is a member of the New World 'justicioid' lineage. |
| Specimen Condition | Live Specimen |
| Copyright |
© 2006
|

| Scientific Name | Metarungia C. Baden |
|---|---|
| Location | Home of K. Balkwill, Johannesberg, South Africa |
| Comments | Metarungia and its sister group Rungia form a clade that is among the paraphyletic group of clades that we refer to as Old World 'justicioids.' |
| Specimen Condition | Live Specimen |
| Copyright |
© 2006
|
About This Page
Rancho Santa Ana Botanic Garden, Claremont, California, USA
Carrie Kiel
Rancho Santa Ana Botanic Garden
Correspondence regarding this page should be directed to Lucinda A. McDade at and Carrie Kiel at
Page copyright © 2009 and Carrie Kiel
All Rights Reserved.
- First online 25 June 2007
- Content changed 25 June 2007
Citing this page:
McDade, Lucinda A. and Carrie Kiel. 2007. Justicieae. Version 25 June 2007 (under construction). http://tolweb.org/Justicieae/52290/2007.06.25 in The Tree of Life Web Project, http://tolweb.org/

