



Septobasidiales
Septobasidiaceae
Daniel Henk


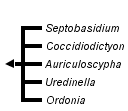

This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
The Septobasidiaceae are primarily defined by their association with scale insects (Coccoidea). Most Septobasidiaceae infect armored scale insects (Diaspididae) then grow out of them to form felty mats on tree branches and leaves. Like their host insects, these fungi can be inconspicuous but are often extremely common in humid tropical, subtropical and mediterranean climates around the world. The genus Septobasidium has been especially interesting to mycologists and nonmycologists because it has a putatively altruistic-mutualistic relationship with its host insects described by J. N. Couch (1938, see below).

Septobasidium septobasidioides growing over a Bryophyte covered branch. The grey layer is the spore bearing surface and it is on top of a brown layer that extends over and among the Bryophytes and along the surface of the branch.
The Septobasidiaceae include at least two different life histories in association with scale insects. In many Septobasidium species, the fungi and insects form a mat of hyphae intertwined with infected and uninfected scale insects. This provides the basis for the hypothesis put forward by Couch in 1931 and expanded in 1938 that some Septobasidium species are actually mutualistic with colonies of scale insects despite renderring infected individuals sterile. In his model, the cost to the colony of sterile infected insects is overcome by uninfected insects covered by the fungus mat being protected from parasitoid wasps and perhaps other environmental stresses. However, in some species of Septobasidium and in each of the other genera in the Septobasidiaceae only a single infected scale insect is covered by an individual fungus or any additional covering is so minor that Couch's model does not apply.
Characteristics

Electron micrograph showing the septal pore in Septobasidium pseudopedicellatum. © and Cindy Henk Socolofsky Microscopy Center Department of Biological Sciences Louisiana State University
The Septobasidiaceae examined to date have simple septum structure with only slight swelling (see above), and relatively few other characters have been noted. Hyphae are clampless, but dikaryotic (two nuclei in each cell) in all species examined to date. Sexually reproducing species form transversely septate basidia giving rise to one to four basidiospores. The number of spores formed is usually consistent within a species. These basidiospores germinate by producing bud-cells that act to infect scale insects via an openning in the cuticle (either an orifice or along a seta). In culture, bud-cells continue to grow as yeasts. In the species where basidiospores have been examined, they have been determined to be haploid and legitimate meiotic products regardless of the number of spores produced from a basidium (Olive 1943). Inside of insects the fungal hyphae differentiate to form haustoria that are often coiled but may take on other defined shapes as well. Hyphae grow out of the insects usually via the anus or vagina, and may fuse with nearby fungal hyphae from other infected insects. Even heavily infected insects remain alive and continue to siphon plant fluid. Hyphae do not grow into living plant cells, and apparently derive nutrition via infected insects. In species that form mats of mycelium, the fungi may produce elaborate pillar-like structures up to several centimeters tall or tiny tents of hyphae that are sometimes called insect traps or houses.
Classification
In Couch's 1938 treatment, the genus Ordonia was subsumed within Septobasidium, but Oberwinkler resurrected the genus in 1989 and added another, Coccidiodictyon. Couch also described the genus Uredinella in 1937 placing it somewhere between rust fungi and Septobasidium. Auriculoscypha was initially described as a member of the Auriculariales, but moved to the Septobasidiales when it was discovered that it was associated with scale insects (Reid and Manimohan 1985; Lalitha and Leelavathy 1990).
Discussion of Phylogenetic Relationships
Molecular analyses support a single origin of the insect association uniting the Septobasidiaceae, but groups within the family lack many unifying characters and relationships among the genera have not been adequately determined (Henk and Vilgalys 2007).
References
Couch, J. N. 1931. The biological relationship between Septobasidium retiforme (B. & C.) Pat. and Aspidiotus osborni New. and Ckll. Quarterly Journal of Microscopical Science 74: 383437.
Couch, J. N. 1937. A new fungus intermediate between the rusts and Septobasidium. Mycologia 29: 665673.
Couch, J. N. 1938. The genus Septobasidium. University of North Carolina Press, Chapel Hill, North Carolina, USA.
Couch, J. N. 1941. A new Uredinella from Ceylon. Mycologia 33: 405410.
Couch, J. N. 1946. Two species of Septobasidium from Mexico with unusual insect houses. Journal of the Elisha Mitchell Scientific Society 62: 8794.
Henk, D.A. and R. Vilgalys. 2007. Molecular phylogeny suggests a single origin of insect symbiosis in the Pucciniomycetes with support for some relationships within the genus Septobasidium. Am J Bot 94: 15151526.
Kumar, A., Celio, G. C., Matheny, P. B., McLaughlin, D. J., Hibbett, D. S., and Manimohan, P. 2007. Phylogenetic relationships of Auriculoscypha based on ultrastructural and molecular studies. Mycological Research 111: 268-274.
Lalitha, C. R. and K. M. Leelavathy. 1990. A coccid association in Auriculoscypha and its taxonomic significance. Mycological Research 94: 571572.
Oberwinkler, F. 1989. Coccidiodictyon, new genus and Ordonia, two genera in the Septobasidiales. Opera Botanica 100: 185192.
Olive, L.S. 1943. Cytology of Various Basidial Types in the Genus Septobasidium. Mycologia 35: 557¬572.
Reid, D. A. and P. Manimohan. 1985. Auriculoscypha, a new genus of Auriculariales (Basidiomycetes) from India. Transactions of the British Mycological Society 85: 532535.
Title Illustrations

| Scientific Name | Septobasidium |
|---|---|
| Location | Florida |
| Specimen Condition | Live Specimen |
| Identified By | Daniel Henk |
| Sex | dikaryon |
| View | Whole colony on Acer sp. |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 2.0. This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 2.0.
|
| Copyright |
©

|

| Scientific Name | Septobasidium gomezii |
|---|---|
| Specimen Condition | Live Specimen |
| Identified By | Daniel Henk |
| Sex | dikaryon |
| Body Part | Probasidia |
| Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 2.0.
|
| Copyright |
©
|
About This Page
The NSF funded 'Deep Hypha' Research Coordination Network (DEB-0090301) facilitated the development of this site. I thank my mother Cindy Henk for providing the electron micrographs and various other types of support.
Imperial College London
Correspondence regarding this page should be directed to Daniel Henk at
Page copyright © 2012
 Page: Tree of Life
Septobasidiales. Septobasidiaceae.
Authored by
Daniel Henk.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Septobasidiales. Septobasidiaceae.
Authored by
Daniel Henk.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 16 August 2005
- Content changed 21 February 2008
Citing this page:
Henk, Daniel. 2008. Septobasidiales. Septobasidiaceae. Version 21 February 2008 (under construction). http://tolweb.org/Septobasidiaceae/51247/2008.02.21 in The Tree of Life Web Project, http://tolweb.org/

