



Dothideomycetes
Conrad Schoch





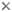
This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
The class Dothideomycetes contains the majority of the fungal species with ascostromatic development and bitunicate asci that were previously placed in the "Loculoascomycetes". The remaining "Loculoascomycetes" are placed in subclass Chaetothyriomycetidae in the diverse order Eurotiomycetes. Historically the majority of these fungi were placed in several different groups under various classifications e. g. “Ascoloculares” (Nannfeldt 1932) “Bitunicatae” (Luttrell 1951) “Loculoascomycetes” (Luttrell 1955) and “Loculoascomycetidae” (Luttrell 1981). A mixture of these names still remain in use in recent literature. Dothideomycetes are often found as pathogens, endophytes or epiphytes of living plants and also as saprobes degrading cellulose and other complex carbohydrates in dead or partially digested plant matter in leaf litter or dung. However their nutritional modes are not limited to associations with plants; several species are lichens, while others occur as parasites on other fungi or members of the kingdom animalia.

Plant pathogenetic Dothideomycetes: Pseudothecia of Cochliobolus heterostrophus on corn leaf seen through a stereo microscope, © Gillian Turgeon
Characteristics

Pyrenophora brizae: Bitunicate asci, one with broken ectotunica. © Robert Shoemaker.
To a casual observer there is little to distinguish the flask, spherical or disk-shaped fruiting bodies of Dothideomycetes from several other groups in the Ascomycota, but they share a distinctive pattern of development. The sexual spore bearing asci develop in pockets (locules) already formed in an unfertilized mass of hyphae (stroma). This is defined as ascolocular development and is in contrast to ascohymenial development found in the majority of other fungal classes.

Guignardia magniferae: Asci with thick layered ectotunica. © Gary Samuels
The double layered, or bitunicate, ascus remains an important character in dothideomycete taxonomy. Bitunicate asci consists of a thick extensible inner layer (endotunica) and a thin inextensible outer layer (ectotunica). Most bitunicate fungi release their ascospores by the extension of the inner ascus wall and the rupture of the outer wall, similar to a "jack-in-the-box" (fissitunicate), but variations are numerous. The broken ectotunica is seen in the picture on the right, and another set of asci with the thick layered ectotunica is shown in the picture on the left.
Another important character, the centrum, is defined as the tissues and cells occupying the cavity of the sexual structure. The hamathecium (Eriksson 1981), i. e. the sterile centrum tissues existing between the asci, is one of the most reliable characters used to delineate ordinal classifications within the Dothideomycetes (see drawings below).

Alternaria alternata: Conidia produced in acropetal chains.
© Keith Seifert
The best studied species in this class tend to be plant pathogens on important agricultural crops. Therefore a large body of work in dothideomycete taxonomy and systematics concerns descriptions of asexual states or anamorphs, the predominant morphological state encountered on agricultural crops. In fact, several families in this class (e. g., Pleosporaceae, Mycosphaerellaceae, Tubeufiaceae) include a high proportion of purely anamorphic species. Many of the anamorphs produce their asexual spores or conidia in chains bourne on specialized hypha, known as conidiophores such as the Alternaria species pictured on the left.

Dothiorella sp.: Conidia borne in pycnidium. © Pedro Crous
In other cases the conidia are within enclosed flask shaped structres (pycnidia) also occuring in several families (e. g., Leptosphaeriaceae, Lophiostomataceae). Pictured to the right is a microscope squash slide sample of conidia from a pycnidium. Other important species include the remainder of the group now informally referred to as the "black yeasts" (the majority of which now belong to the Eurotiomycetes) characterized by the production of dark, slimy colonies and sporulation patterns that resemble the budding of true yeasts, but are actually reduced versions of cells bearing the conidia directly.
Several classification systems for Dothideomycetes exist but the most important concepts revolves around three different developmental types originally exemplified by the genera Dothidea, Pleospora and Elsinoё (Luttrell 1951, 1955) and later extended (e. g. Barr, 1987). These species form part of the orders, Dothideales, Pleosporales and Myriangiales. The presence of pseudoparaphyses (sterile cells extending down from the upper portion of the sexual structures, initially attached at both ends, although the upper part may become free) is a notable character for the Pleosporales, together with mainly ostiolate flask shaped sexual structures. Conversely, the absence of pseudoparaphyses and the presence of groups of asci are important in the Dothideales. The Myriangiales also do not have pseudoparaphyses, but produce single asci in multiple locules. Several additional orders, defined by combining centrum and ascomal characters, form part of the most recent classification proposed by Eriksson (2006).



1. Dothidea sambuci, multiascus locules typical of the Dothidea type centrum. 2. Elsinoё canavaliae: Single ascus locules typical of the Elsinoё type centrum. 3. Pleospora herbarum, several asci in a flask a shaped sexual structure (pseudothecium) with pseudoparaphyses exemplifying the Pleospora type centrum. Drawings from Von Arx and Müller (1974), © The Centraalbureau voor Schimmelcultures.
Discussion of Phylogenetic Relationships
The use of molecular data, mainly as DNA sequences obtained from the ribosomal RNA genes, has added additional challenges for mycologists trying to describe a defining morphology for the Dothideomycetes. A group of fungi with bitunicate asci and ascolocular development, broadly known as "black yeasts" were moved from this class to the Eurotiomycetes and the subclass Chaetothyriomycetidae, mainly based on DNA sequence data. Some species in this informal group are still retained in the Dothideomycetes however. This emphasizes the fact that a number of morphological characters in these groups are either due to the retention of ancestral characters, or convergent evolution. The large number of genera that remain uncertainly classified in either Dothideomycetes or Chaetothyriomycetidae is a testament to this.
The latest phylogenetic evidence (Schoch et al. 2006) finds support for at least two large groups (newly designated as subclasses) correlating with the three orders mentioned above. The two orders without pseudoparaphyses (Dothideales and Myriangiales) were shown to be related in Dothideomycetidae, while the large order with pseudoparaphyses (Pleosporales) formed most of Pleosporomycetidae. This builds on pioneering molecular studies done by several authors before (e. g. Berbee, 1996; Liew et al 2000; Lindemuth et al 2001). Several orders and groups remain unplaced, and several more do not have representative DNA sequences. For example, a number of lichen lineages in the Trypetheliaceae previously thought to be related to groups in the Eurotiomycetes are now placed in Dothideomycetes based on molecular data (Del Prado et al. 2006). It therefore seems clear that this part of the tree of life will remain quite dynamic for the foreseeable future.
References
Arx von J, Müller E. 1975. A re-evaluation of the bitunicate ascomycetes with keys to families and genera. Studies in Mycology 9:1-159.
Barr ME. 1987. Prodromus to class Loculoascomycetes. Amherst, Mass.: M.E. Barr Bigelow. 168 p.
Berbee ML. 1996. Loculoascomycete origins and evolution of filamentous ascomycete morphology based on 18S rRNA gene sequence data. Molecular Biology and Evolution 13:462-470.
Del Prado RI, Schmitt I, Kautz S, Palice R, Lücking R, Lumbsch HT. 2006. Morphological and molecular evidence place the Tryphetheliaceae in the Dothideomycetes. Mycological Research 110: 511-520.
Eriksson OE. 1981. The families of bitunicate ascomycetes. Opera Botanica 60:1-220.
Liew, E. C., Aptroot, A. and Hyde, K. D. 2000. Phylogenetic significance of the pseudoparaphyses in Loculoascomycete taxonomy. Molecular Phylogenetics and Evolution 16: 392-402.
Lindemuth R, Wirtz N, Lumbsch HT. 2001. Phylogenetic analysis of nuclear and mitochondrial rDNA sequences supports the view that loculoascomycetes (Ascomycota) are not monophyletic. Mycological Research 105:1176-1181.
Nannfeldt JA. 1932. Studien über die Morphologie und Systematik der nicht-lichenisierten inoperculaten Discomyceten. Nova Acta Regiae Societatia Scientiarum upsalensis. (Inaugural dissertation) Uppsala: Almqvist & Wiksells. 368p.
Luttrell ES. 1951. Taxonomy of Pyrenomycetes. University of Missouri Studies in Sciences. 24.
Luttrell ES. 1955. The ascostromatic Ascomycetes. Mycologia 47:511-532.
Luttrell ES. 1981. The Pyrenomycete centrumLoculoascomycetes. In: Reynolds DR, editor. Ascomycete Systematics: The Luttrellian Concept. New York: Springer-Verlag. p 124137.
Schoch CL, Shoemaker RA, Seifert KA, Hambleton S, Spatafora JW, Crous PW. 2006.A multigene phylogeny of the Dothideomycetes using four nuclear loci. 2006. Mycologia 98:1043-1054
Title Illustrations

| Scientific Name | Hysteropatella prostii |
|---|---|
| Location | Germany, Baden-Württemberg, Tübingen, Pfrondorf |
| Comments | Hysterothecia of Hysteropatella prostii on living trunk of Malus domestica |
| Specimen Condition | Live Specimen |
| Identified By | H.-O. Baral |
| Life Cycle Stage | Teleomorph |
| Body Part | Ascoma |
| Copyright | © |

| Scientific Name | Cucurbitaria laburni |
|---|---|
| Location | Austria, Wien 12, Schömbrunn, "Gloriette, Weg zur Hohenbergstraße" |
| Comments | Pseudothecia of Cucurbitaria laburni on branch of Laburnum anagyroides |
| Specimen Condition | Live Specimen |
| Identified By | Walter Jaklitsch |
| Life Cycle Stage | Teleomorph |
| Body Part | Ascoma |
| Copyright | © |

| Scientific Name | Stylodothis puccinioides |
|---|---|
| Comments | Multiascus locules |
| Specimen Condition | Live Specimen |
| Identified By | Robert Shoemaker |
| Sex | Teleomorph |
| Copyright | © |
About This Page

Oregon State University, Corvallis, Oregon, USA
Correspondence regarding this page should be directed to Conrad Schoch at
Page copyright © 2007
 Page: Tree of Life
Dothideomycetes.
Authored by
Conrad Schoch.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Dothideomycetes.
Authored by
Conrad Schoch.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 19 March 2007
- Content changed 19 March 2007
Citing this page:
Schoch, Conrad. 2007. Dothideomycetes. Version 19 March 2007 (under construction). http://tolweb.org/Dothideomycetes/29051/2007.03.19 in The Tree of Life Web Project, http://tolweb.org/

